Екатеринбург: УрО РАН, 2009. 100с.
Род Слепушонки - Ellobius Fischer, 1814
Особенности морфологии некоторых черепных структур и жевательной мускулатуры,
сближающие их с Cricetinae, дали основание некоторым исследователям вывести слепушонок за рамки группы Arvicolinae
(Громов, Поляков, 1976;
Repenning, 1968). Но результаты изучения особенностей ранних (плиоценовых) этапов эволюции Ellobiusini (Топачевский, Рековец, 1982)
показали их несомненно большую близость полевкам, чем настоящим хомякам.
Плиоценовые формы слепушонок (Топачевский,1965; 1973;
Топачевский, Рековец, 1982)
имеют более "полевочий" облик зубов (более выраженные входящие углы, более заостренные вершины выходящих углов,
более сложный рисунок жевательной поверхности зубов, особенно третьих).
Более простой рисунок жевательной поверхности современных слепушонок следует рассматривать как вторичное явление
(Тесаков А.С., устное сообщение; Якименко, Воронцов 1982). Оно может быть связано с тем, что гипсодонтность моляров увеличивалась за счет базальной
части коронки и, как следствие, относительного сокращения онтогенетических стадий апикальной части зуба.
род представлен одним видом –
Cлепушонка обыкновенная - Ellobius (Ellobius) talpinus Pallas, 1770.
Для позднеплейстоценовой фауны Южного Зауралья А.Г. Малеевой (Малеева,1982; Малеева, Стефановский, 1988) отмечено нахождение Ellobius ex gr. tancrei Blas., 1884 (местонахождение Верхняя Алабуга, первая половина позднего плейстоцена).
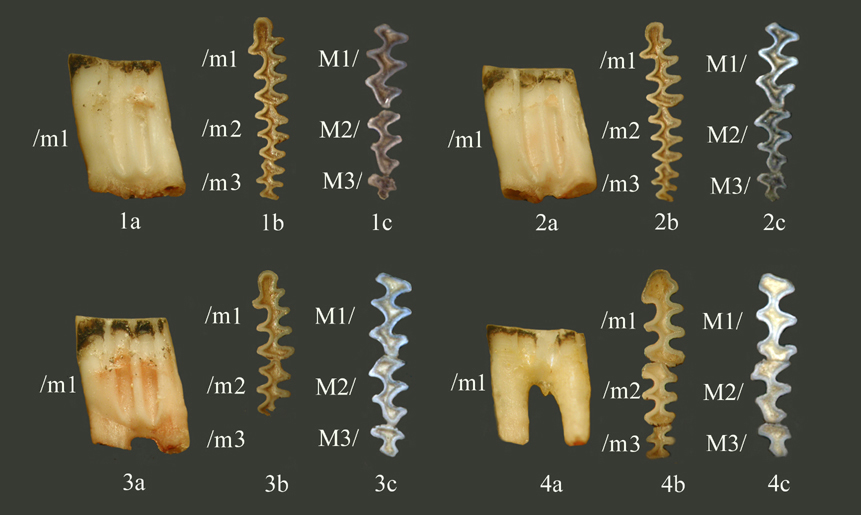
Характеристика зубной системы
Верхние и нижние резцы направлены вперед и сильно выступают из ротовой полости.
Положение нижнего резца диагональное: корневые отделы m/1 и m/2 лежат снаружи, а m/3 обычно над альвеолярной частью нижнего резца.
В отличие от прочих рассматриваемых видов полевок, альвеолярный отросток нижнечелюстной кости принимает вид обособленного выроста (processus alveolaris - как у слепышей) и может достигать уровня конца сочленовного и даже заходить за него (Громов, Поляков, 1977).
Верхние резцы относительно слабо изогнуты по сравнению с остальными полевками; их основания внутри челюсти заходят назад за передние основания M1/ (Огнев, 1948).
Форма резцов (как верхних так и нижних) связана с приспособлением слепушонок к рытью резцами (Зубцова, 1978).
Размеры коренных зубов современных Ellobius talpinus, мм: (min-max, n=40, Челябинская область):
Длина: М1/: 2,6-3,25; М2/: 2,0-2,5; М3/: 0,95-1,3; /m1: 2,8-3,35; /m2: 1,85-2,25; /m3: 1,3-1,7.
Ширина: М1/: 1,2-1,6; М2/: 1,15-1,5; М3/: 1,0-1,3; /m1: 1,15-1,4; /m2: 1,1-1,35; /m3: 0,8-1,1.
Дентиновые поля призм щечных зубов широко слиты на жевательной поверхности, парные призмы смещены относительно друг друга не значительно. Входящие углы округлые, что усиливается при значительной стертости коронки.
Отложения наружного цемента во входящих углах - нет.
Эмаль толстая, не дифференцированная по толщине.
Призматическая складка может формироваться на передне-буккальной стороне антероконидного отдела /m1. В.А. Топачевский (1965) предложил этот признак как диагностический для E. tancrei (100%, против 10% у обыкновеной слепушонки), но по уточненным данным Л.В. Якименко и Н.Н. Воронцова (1982) встречаемость этого признака в отдельных популяциях E. talpinus может достигать 30-57% (выборки из Челябинской и Омской областей) , в то время как среди типичных E. tancrei в отдельных популяциях этот признак отмечен только на уровне 3,6% (окрестности оз. Исык-Куль), 17% (Тува, Монголия). Для большинства популяций E. tancrei этот признак действительно выражен у 100% особей (Якименко, Воронцов 1982). Необходим более детальный анализ выраженности этого признака в зависимости от онтогенетической стадии и степени стертости коронки зуба.
Дентиновые тракты очень низкие и могут доходить до жевательной поверхности только на сильно стертых коронках.
Корни у щечных зубов М1/, М2/, /m1, /m2 имеют по два корня (иногда имеется костная пластинка, приросшая к переднему корню), М3/ и /m3 имеют по одному корню. На передних зубах корни формируются раньше. Максимальная длина изолированных корней /m1 не превышает 3,5 мм (Евдокимов, 1997). Выраженность корней может быть использована для определения индивидуального возраста животного.
С.В. Кириков (1952) выделил пять возрастных групп по степени выраженности корней /m1:
1) juv. – корней нет (коронка не сформирована, призмы внизу не замкнуты); 2) subad. – корней нет или они только начали формироваться (коронка сформирована); 3) ad.1 - длина корней не превышает ¼ общей высоты зуба; 4) ad.2 - длина корней от ¼ до ½ общей высоты зуба; 5) seniles - длина корней более ½ высоты зуба.
Н.Г. Евдокимов (1997; 2001) предложил метод определения календарного возраста слепушонок по степени выраженности корней:
у сеголеток с марта до октября корни увеличиваются от 0-0,1 до 0,7 мм; у однолеток с марта до октября от 0,2-0,9 до 0,7-1,3 мм; у двухлеток за тот же период – от 1,0-1.6 до 1,5-2,1 мм; трехлеток – от 1,7-2,2 в марте до 2,3-2,9 в октябре; четырехлеток – от 2,3-2,7 в марте до 2,7-3,1 в октябре; пятилеток –от 2,8-3,1 в марте до 3,6 в октябре; шестилеток от 3,2-3,4 в марте до полного стачивания коронки (Евдокимов, 1997).
Динамика роста корней имеет ступенчатый характер: осенью рост корней замедляется, а во время зимовки прекращается совсем. Этот факт трактуется Н.Г. Евдокимовым как еще одно косвенное подтверждение зимней спячки
E. talpinus. С возрастом темп роста корней замедляется (Евдокимов, 1997; 2001; 2002).
Строение и морфотипические характеристики жевательной поверхности щечных зубов:
M1/ – передняя непарная лопасть, Т1, Т2, Т3, Т4;
M2/ – передняя непарная лопасть, Т2, Т3, Т4;
М3/ – упрощенного строения, значительно укорочен, по сравнению с одноименным зубом зеленоядных
форм;
/m1 – задняя непарная лопасть, Т1, Т2, Т3 и первично простой антероконид в форме трилистника у молодых животных и без выраженных входящих углов у старых;
/m2 - задняя непарная лопасть, Т1, Т2, Т3, Т4;
/m3 - задняя непарная лопасть, Т1, Т2, Т3 и слабо выраженный Т4.
Изменчивость характеристик жевательной поверхности. Изменчивость конфигурации жевательной поверхности зубов прежде всего связана с стадией (степенью) сформированности (сточенности) коронки. Так, антероконид /m1 у молодых особей имеет вид трилистника, соответствующие ему выступающие углы хорошо развиты, однако они доходят лишь до ½ коронки, в результате чего антероконид у особей со значительно стертыми зубами упрощается, имеет округлую форму. Треугольники жевательной поверхности /m1 у молодых животных образуют единое пространство, которое может быть отделено, а может быть слито с антероконидом. У старых Т3 полностью слит с антероконидом. Кроме того, зубы с нестертой коронкой имеют жевательную поверхность, на которой входящие углы глубже, несколько острее, чем на стертых зубах. У старых особей входящие углы отчетливо округлые. Жевательная поверхность упрощается по мере приближения к шейке, может наступить полное стирание коронки, причем в первую очередь стираются /m3 и /m2.