Екатеринбург: УрО РАН, 2009. 100с.
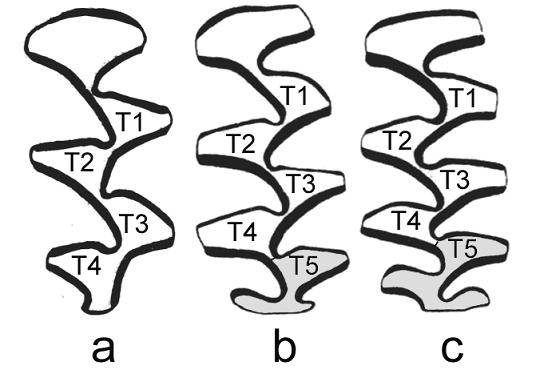
Жевательная поверхность Praedicrostonyx compitalis Zazhigin, 1974 (a) и наиболее усложненные варианты жевательной поверхности Dicrostonyx torquatus (b, c).
Подходы к оценке изменчивости и таксономической значимости морфотипических характеристик коренных зубов были предложены А.К. Агаджаняном (1973, 1976) и в дальнейшем развиты В.С. Зажигиным (1976, 2003), В.А. Кочевым (1983, 1993), Н.Г. Смирновым (Смирнов, Большаков, 1985; Смирнов, Большаков, Бородин, 1986; Смирнов и др., 1997). В основе таксономической диагностики копытных леммингов по коренным зубам лежит оценка соотношения архаичных и «прогрессивных» морфотипов зубов. Для копытных леммингов плейстоцена Н.Г. Смирновым (Смирнов, Большаков, Бородин, 1986) была предложена классификация морфотипов коренных зубов, основанная на качественной оценке дополнительных элементов.
(по Смирнов, Большаков, Бородин, 1986).
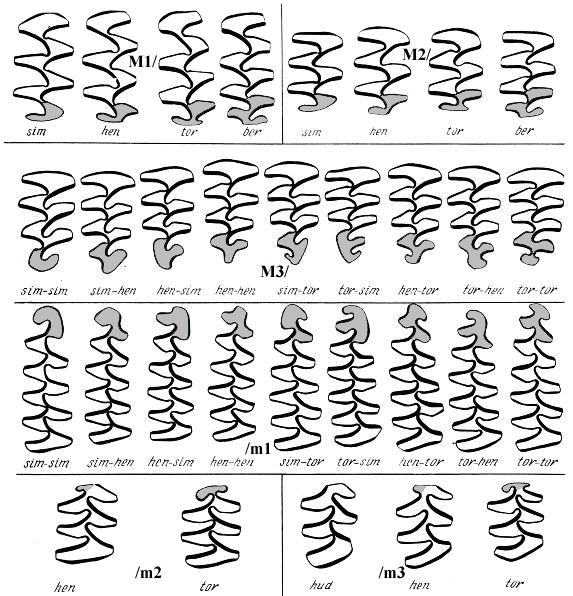
M1/ - первый верхний зуб, M2/ - второй верхний зуб, M3/ - третий верхний зуб, /m1 - первый нижний зуб, /m2 - первый нижний зуб, /m3 - первый нижний зуб.
Обозначения морфотипов: sim - "simplicior", hud - "hudsonius", hen - "henseli", tor - "torquatus", ber - "beringius". Морфотипы m/1 и M3/ определяются по выраженности входящих углов с буккальной и с лингвальной сторон
(по Смирнов, Большаков, Бородин, 1986)
Для оценки степени развития дополнительных элементов М1/ и М2/ возможно использование и количественных (метрических) показателей. Для количественной оценки степени усложненности в пределах морфотипа "simplicior" Н.Г. Смирнов (Смирнов и др., 1997) предложил использовать индекс a/b, где а - ширина дополнительной призмы, b- ширина предшествующей призмы
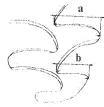
Схема измерений ширины треугольников на первом и втором верхних молярах, применяемая для определения стадии развития дополнительного элемента в пределах морфотипа simplicior (по Смирнов и др., 1997 с. 76)
Цифровое выражение степени развития получалось при делении ширины «b» на ширину «а». Это отношение равно единице, когда треугольники имеют равную ширину, и оно тем меньше, чем менее развит новый треугольник. В числовом выражении, по данным Н.Г. Смирнова (Смирнов и др., 1997), соответствующие значениям индекса a/b выделяется четыре стадии: s1 (a/b=0,59 и менее); s2 (a/b=0,6-0,69); s3 (a/b=0,7-0,79); s4 (a/b=0,8 и более).
Для количественной оценки выраженности входящего угла на дополнительной призме используется методика В.А. Кочева, основанная на измерении угла при помощи окуляр-транспортира (Кочев, 1983; Смирнов, Большаков, Бородин, 1986; Смирнов и др., 1997)
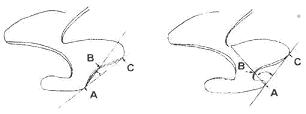
Схема измерения угла ВАС для оценки степени развития дополнительных элементов на М1/ и М2/ в пределах морфотипов henseli и torquatus (по Смирнов и др., 1997 с. 77)
Величина угла ВАС измерялась с точностью до 5 градусов, максимальные значения – 120 градусов. В этом случае морфотипы М1/ и М2/ выделяются по следующим критериям:
- морфотип simplicior – 0 градусов – делится на 4 класса (s1, s2, s3 и s4) с соотетствующими значениями индекса а/b (0,59 и менее; 0,6-0,69; 0,7-0,79; 0,8 и более),, по Н.Г. Смирнову (Смирнов и др., 1997);
- морфотип henseli делится на 5 классов (h1 - h5) с соответствующими значениями углов: 5 и 10°, 15 и 20°, 25 и 30°, 35 и 40°, 45 и 50°.
- морфотип torquatus может быть представлен шестью классами (t1 - t6), соответственно 55 и 60°; 65 и 70°; 75 и 80°; 85 и 90°; 95 и 100°; 105 и более градусов.
Как крайний случай (недостаточный объем выборок) по мнению Н.Г. Смирнова возможно ограничиться выделением трех морфотипов (simplicior, henseli, torquatus) по крайним значениям признаков (Смирнов и др., 1997).
В течение плейстоцена на рассматриваемой территории были представлены следующие виды: Dicrostonyx renidens Zazhigin 1974, D. simplicior Fejfar 1966, D. ocaensis Alexandrova 1982 (Александрова, 1982), D. henseli Hinton 1910, D. gulielmi Sandford 1870, D. torquatus. Виды приведены в последовательности от наиболее древнего (D. renidens) к современному копытному леммингу (D. torquatus).
Таксономический статус и валидность D. ocaensis, D. henseli, и D. gulielmi по разному оцениваются отдельными исследователями. К. Ковальский считает, что D. henseli, и D. gulielmi могут быть сведены к синонимам (Sutcliffe, Kowalski, 1976). Этой же точки зрения придерживаются А. Надаховский (Nadachowski 1982), А.К. Маркова (Markova 2006), Н.Г. Смирнов (Смирнов и др., 1997) и др., отрицающие таксономическую самостоятельность D. henseli.
По мнению А.К. Агаджаняна (Агаджанян, Глушанкова, 1986 ), морфотипические характеристики моляров D. ocaensis соответствует подвидовому рангу - D. simplicior ocaensis.
В.С.Зажигин (2003) считает выделение самостоятельной таксономической группы «ocaensis» не правомерным, поскольку оно не подтверждено имеющимся материалом.
Кроме того, В.С. Зажигин считает, что D. gulielmi= D.torquatus, напоминая, что еще М. Хинтон показал идентичность морфологических характеристик моляров этих двух видов. Исходя из объема вида, обозначеннного М. Хинтоном для D. henseli, "следует все известные среднеплейстоценовые остатки копытных леммингов Русской равнины относить к D. henseli. До сих пор разные исследователи остатки этого вида определяли как D. simplicior, D. cf. simplicior, D. ex. gr. simplicior, D. aff. gulielmi" (цит. по Зажигин, 2003, с.17-18). В той же работе отмечается, что это относится и к копытным леммингам среднего плейстоцена Западной Сибири.
Все выше перечисленные авторы указывают на то, что при выделении таксона внутри рода Dicrostonyx нельзя ограничиваться морфотипическими характеристиками одного зуба.
ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВОГО СТАТУСА копытных леммингов Н.Г. Смирнов предлагает метод типологизации М1/ и М2/ по морфотипам (Смирнов и др., 1997). Типология зубных систем основана на отнесении выборок к одному из трех известных видов: D. simplicior, D. gulielmi, D. torquatus, в тех случаях когда типичный для данного вида морфотип преобладает в выборке как на М1/, так и на М2/. В пределах каждого вида выделяются по три морфы по степени доминирования более простых и более сложных морфотипов на первых и вторых зубах. Кроме видовых форм Н.Г. Смирнов предлагает выделять, используя открытую номенклатуру, переходные, название которых состоит из двух видовых (на первом месте название вида по преобладающему морфотипу) (Смирнов и др., 1997).
По мнению Н.Г. Смирнова (Смирнов, Кузьмина, Коурова, 1999; Смирнов, Федоров, 2003), эволюция евразийских копытных леммингов осуществлялась по модели прерывистого равновесия, и переход от D. guilielmi к D. torquatus произошел в интервале 15-13 тысяч лет, что демонстрируется скачкообразным увеличением скорости морфологических изменений зубной системы.
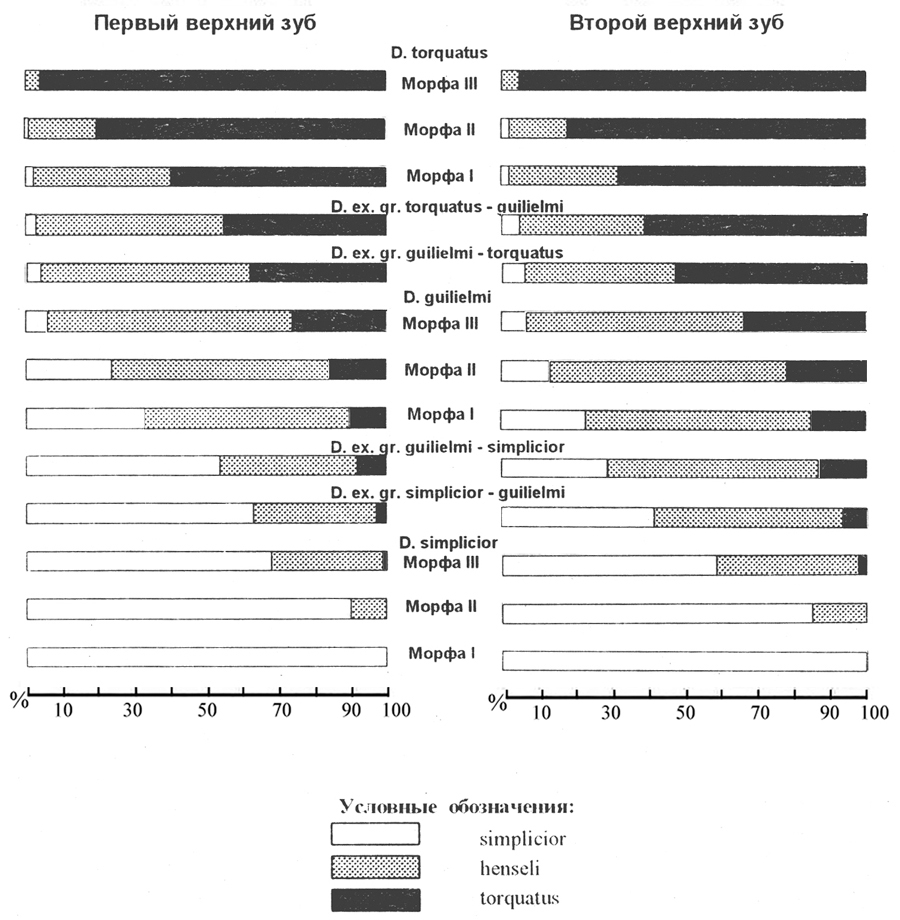
Соотношение морфотипов simplicior, henseli, torquatus в разных таксонах копытных леммингов (по Н.Г. Смирнову (Смирнов и др., 1997)
Иной взгляд на таксономию плейстоценовых копытных леммингов предлагает В.С. Зажигин (2003), хотя в основе лежит тот же принцип анализа частот морфотипов, что и в рассмотренных выше: в линии D. simplicior - D. torquatus каждый вид имеет несколько морфотипов M1/ и M2/. На основании степени усложненности коренных зубов В.С. Зажигин выделяет следующие филогенетически связанные виды:
- Dicrostonyx renidens Zazhigin - наличие на М1/ и М2/ двух небольших округлых призм, ни одна из которых не достигает формы треугольника; на /m2 зачатки дополнительного образования на передней призме. Время существования - конец эпохи Матуяма - начало Брюнес; руководящая форма поздней стадии олерского комплекса млекопитающих Колымской низменности (Шер и др., 1979; Зажигин 2003).
- D. simplicior Fejfar - на М1/ и М2/ дополнительная оформившаяся призма; изменчивость М2/ допускает отдельные экземпляры с морфотипом "henseli"; морфотипическая изменчивость /m2 очень широка - от "simplicior" до "torquatus. В окскую ледниковую эпоху у D. simplicior отмечен один морфотип М1/ - "simplicior" и два морфотипа М2/ - "simplicior" и "henseli". Время появления - ранний неоплейстоцен; вид широко распространен в средних широтах Евразии (Fejfar, 1966; Heller, 1968; Kowalski, 1977; Nadachowski 1982; Агаджанян, Глушанкова 1986; Смирнов, Большаков, Бородин, 1986 и др.).
- D. henseli Hinton - главное отличие от D. simplicior - наличие на /m3 D. henseli одного или двух дополнительных элементов на передней призме этого зуба и более сложные морфотипы М1/, и М2/: в днепровское время у М1/ и М2/ два морфотипа - "simplicior" и "henseli", а в позднеднемосковское время три морфотипа - "simplicior", "henseli" и "torquatus".
- D. gulielmi Sanford = D. torquatus Pallas - поздний плейстоцен - современность.
Изменчивость современного D. torquatus описывается четырьмя морфотипами - "simplicior", "henseli", "torquatus" и "beringius".
Наличие среди современных представителей рода Dicrostonyx таких видов, находящихся на разной стадии усложнения жевательной поверхности моляров, как D. torquatus и D. hudsonius, может свидетельствовать, как мы уже отмечали ранее (Смирнов, Большаков, Бородин, 1986), о том что и в течение плейстоцена могла существовать такая же ситуация, то есть в пределах плейстоценового ареала рода копытных леммингов одновременно могли существовать виды, различающиеся в масштабе по крайней мере одной эволюционной ступени.