Екатеринбург: УрО РАН, 2009. 100с.
На территории Урала и Западно-Сибирской низменности в настоящее время представлена двумя подвидами, различающимися в том числе и по размерам: M. (S.) g. major Ognev, 1918 и M. (S.) g. gregalis Pallas.
При оценке эволюционного уровня узкочерепных полевок можно использовать как количественные показатели – выраженность буккального угла передней непарной петли (Смирнов, Большаков, Бородин, 1986), так и качественные – соотношение морфотипов (Большаков, Васильева, Малеева, 1980). Схема морфологических преобразований вида достаточно подробно изучена на всей территории Евразии.

A - верхний зубной ряд, В - нижний зубной ряд.
 |
|
|
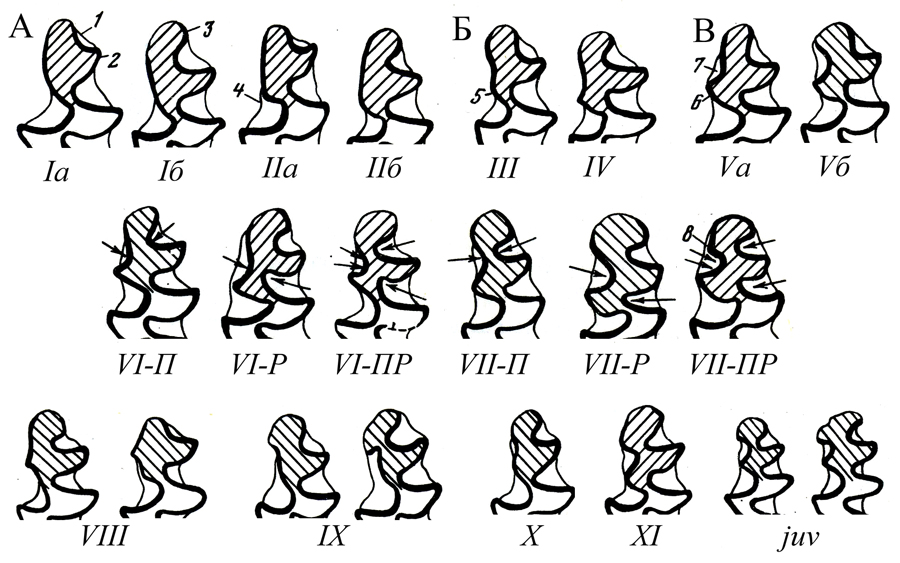
Основные морфотипы /m1 позднеплейстоценовой и современной узкочерепной полевки по А.Г.Малеевой (Большаков, Васильева, Малеева, 1980):
А - грегалоидные (I, II); Б - грегало-микротидные (III, IV); В - микротидные. Стрелки характеризуют тенденцию к разделению эмалевых полей для усложненных вариаций VI и VII морфотипов (П - питимисный, Р - ратицепоидный, ПР - смешанный). VIII-XI - резервные морфотипы |
|
 |
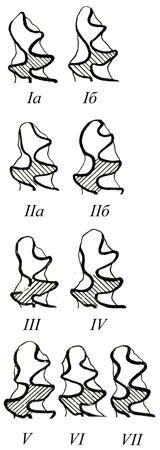 |
| Питимисные вариации основных морфотипов /m1 узкочерепной полевки по А.Г. Малеевой (Малеева, 1977; Малеева, Шувалова, 1980; Большаков, Васильева, Малеева, 1980) | |
 |
|
|
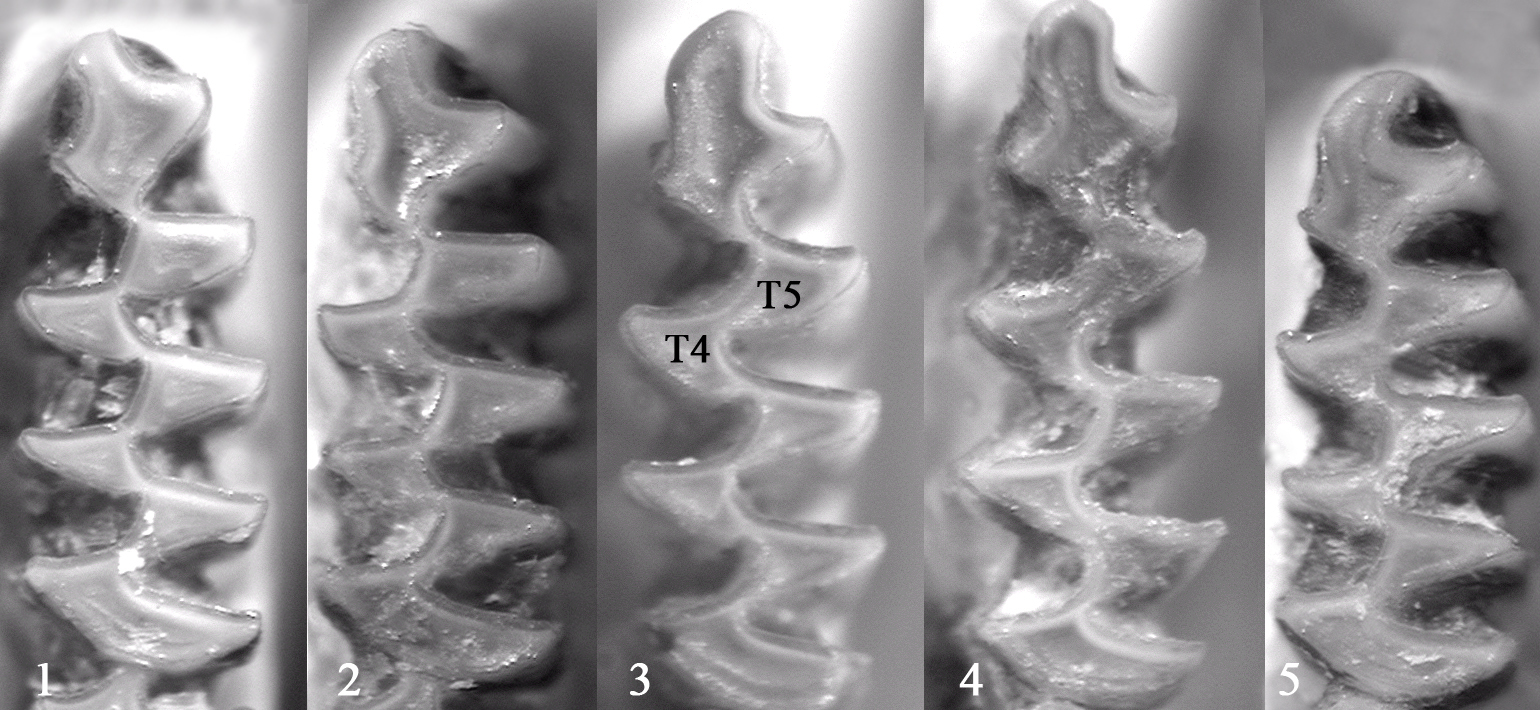
Некоторые варианты формы передней непарной петли и слияния дентиновых полей T4-T5 современных M. g. major на зкземплярах 3 и 4 дентиновые поля Т4 и Т5 слиты (питимисное слияние) |
|
 |
|
|
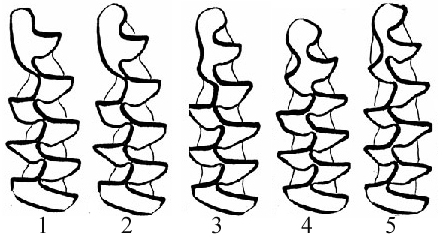
Основные морфотипы /m1 узкочерепной полевки по А.В. Бородину (Смирнов, Большаков, Бородин, 1986): морфотип I - 1,2; морфотип II - 3; морфотип III - 4, 5. |
|
|
Все многообразие морфотипов узкочерепных полевок можно, на мой взгляд, свести к трем группам: |
|
 |
|
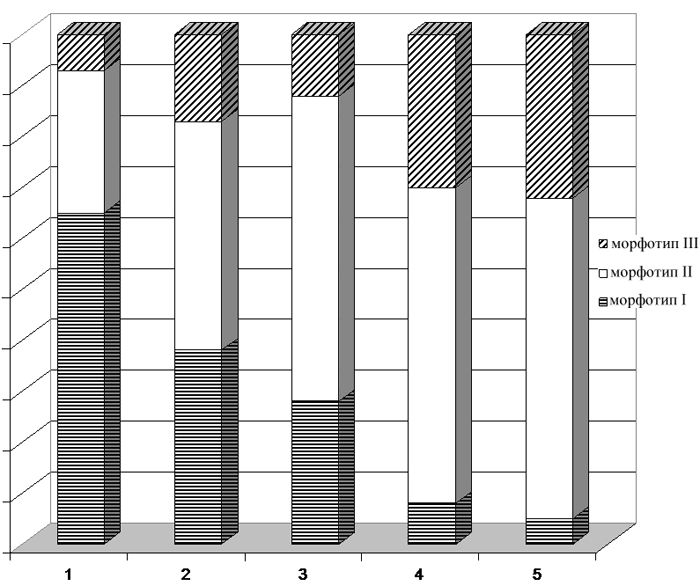
| Соотношение морфотипов /m1 M. gregalis из ископаемых и современных выборок Западно-Сибирской равнины. Выборки из местонахождений: 1 – Чембакчино (средний неоплейстоцен), 2 - Ярсино (рубеж среднего и позднего неоплейстоцена), 3 – 430 км (конец позднего неоплейстоцена), 4 – отловы современных M. gregalis major (Южный Ямал), 5 - отловы современных M. gregalis gregalis (Курганская обл.) | |
|
Все экземпляры первых нижних коренных зубов, отнесенные
к этому виду, не выходят за границы размерных и морфотипических
характеристик, описанных для М. gregalis Pallas (Агаджанян, 1971; Большаков, Васильева, Малеева, 1980;
Головачев и др., 2001; Громов, Поляков, 1977; Дупал, 1998,
2000а, 2000б; Зажигин, 1980; Малеева, 1977;
Смирнов и др., 1990; Кочев, 1991; Мотузко, 1992;
Малеева, Шувалова, 1980; Маркова, 1986; Рековец, 1978, 1985,
1994; Смирнов, 1996; Тютькова, 1991;
Nadachowski, 1982; Válóczi, 1999;и др.). Изменение во времени соотношения трех типов передней непарной петли антероконидного отдела узкочерепной полевки на территории Западно-Сибирской низменности
не противоречит тенденциям, описанным для узкочерепной полевки в других регионах
(Большаков, Васильева, Малеева, 1980; Малеева, 1977; Nadachowski, 1982 и др.).
Наибольший процент первой (морфотип I), "архаичной", группы приходится на среднеплейстоценовых узкочерепных полевок из местонахождений Чембакчино и Семейка;
в местонахождении Ярсино (начало позднего плейстоцена) при значительной доле первой группы морфотипов уже преобладает II группа "типичных" морфотипов
и увеличивается доля III группы. В позднеплейстоценовых местонахождениях из низовьев Оби доля второй группы еще больше возрастает и практически
достигает уровня, характерного современным популяциям как северного, так и южного подвида. Таким образом, степень сложности антероконидного отдела
/m1 может использоваться для определения относительного геологического возраста вмещающих пород (Большаков, Васильева, Малеева, 1980;
Маркова 1982; Смирнов, Большаков, Бородин, 1986; Рековец, 1994 и др.). |
|
|
Вопросы подвидовой систематики и закономерностей географической изменчивости узкочерепной полевки на всем ее ареале рассматривался рядом авторов (Дупал, 1998; 2000а; 2000б;
Голенищев, Петровская, 2002 и др.). В результате проведенных работ не выявлен половой диморфизм по морфологическим признакам.
Возрастная изменчивость проявляется в быстром росте до 3 месяцев и более замедленном - до 7 месяцев. |
|
|
Происхождение современных подвидов узкочерепной полевки Урала и Западно-Сибирской низменности. Все плейстоценовые местонахождения нижнего Прииртышья и Приобья находятся вне ареалов современных подвидов узкочерепной полевки,
поэтому интересен вопрос о "родственных" отношениях ископаемых M. gregalis с современными северными и южными западносибирскими
подвидами узкочерепной полевки. Так, А.Г. Малеева (Большаков, Васильева, Малеева, 1980; Малеева, 1983) считает,
что в течение неоплейстоцена в Среднем Зауралье обитали на одних территориях две формы узкочерепной полевки - крупная и мелкая.
Первая из них мигрировала в северном направлении и явилась предковой формой северного подвида, а вторая, отступая в южном направлении,
дала начало современному южному подвиду. |
|
| Основное меню подсемейства Arvicolinae | |