Екатеринбург: УрО РАН, 2009. 100с.
В позднем плейстоцене и голоцене представлены одним видом:
пеструшка желтая - E. luteus Eversm.
| Желтая пеструшка наиболее близка пеструшке степной, от которой отличается более крупными размерами и более простым строением M3/ и /m1 | ||
 |
 |
Морфотипы жевательной поверхности /m1 позднеплейстоценовых E. luteus Южного Зауралья по А.Г. Малеевой (Малеева, Филева, 1979). |
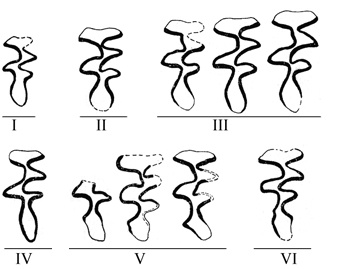 |
Морфотипы жевательной поверхности M3/ позднеплейстоценовых E. luteus Южного Зауралья по А.Г. Малеевой (Малеева, Филева, 1979) | |
|
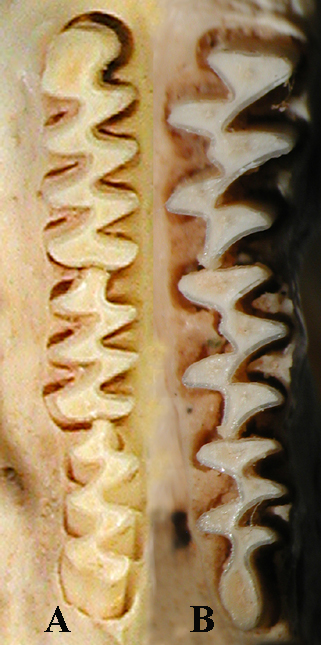
A - нижний зубной ряд В - верхний зубной ряд |
||
Размеры коренных зубов /m1 современных
E. luteus Зайсанской котловины (мм, min-a-max):
Длина 2,85-3,17-3,8; ширина 1,00-1,17-1,35
):
Положение нижнего резца диагональное: корневые отделы /m1 и /m2 лежат снаружи, а /m3 с лингвальной стороны нижнего резца.
Дентиновые поля призм щечных зубов изолированы, но могут быть частично слиты (ширина слияний дентиновых полей основных призм /m1 больше толщины эмали).
Отложения наружного цемента во входящих углах отсутствуют.
Эмаль дифференцирована по микротусному типу: более тонкая на выпуклых стенках призм, чем на вогнутых и всегда тоньше во вершинах входящих углов.
Дентиновые тракты на передне-буккальной стороне головки непарной петли /m1, задней непарной петле М3/, по обеим сторонам непарных лопастей щечных зубов, а также на вершине LSA2 М1/. У молодых животных эмаль на жевательной поверхности может быть непрерывна.
Корни у щечных зубов не формируются.
Строение и морфотипические характеристики жевательной поверхности щечных зубов:
М1/-М2/ без добавочных петель, на внутренних стенках призм Т2 этих зубов имеется складка эмали.
Морфотипы моляров рода Eolagurus
(кроме M3/) могут совпадать с морфотипами рода Lagurus: для /m1 - наиболее простые морфотипы
L.lagurus и наиболее усложненные морфотипы E.luteus
(Агаджанян, Маркова, 1984; Малеева, 1978; Малеева, Филева, 1979).
M1/ – передняя непарная лопасть, Т1, Т2, Т3, Т4; на внутренней стенке призм Т2 имеется складка эмали;
M2/ – передняя непарная лопасть, Т2, Т3, Т4; на внутренней стенке призм Т2 имеется складка эмали;
М3/ - передняя непарная лопасть, Т2, Т3, Т4, задняя непарная петля. Дентиновое поле задней непарной петли чаще всего слито с дентиновым полем Т4. На лингвальной стороне Т2 может формироваться лагурусная складка, как на M1/ и M2/. Форма задней непарной петли от практически круглой до вытянутой овальной, на лингвальной стороне может формироваться призматическая складка с дентиновым трактом. Хорошо выраженная складка эмали может быть в вершине первого входящего угла (Малеева, Филева, 1979; Яковлев, 1996).
/m1 – задняя непарная лопасть, Т1, Т2, Т3, антероконид вторично усложнен: формируются T4, T5; дентиновые слияния между отделами антероконида и основными дентиновыми призмами могут быть существенно больше толщины эмали, передняя непарная петля отделена имеет округлую, округло-трапециевидную форму, на буккальной стороне может формироваться входящий угол (Малеева, Филева, 1979; Тропин, 1983; Яковлев, 1996).
/m2 - задняя непарная лопасть, Т1, Т2, Т3, T4;
/m3 – задняя непарная лопасть, Т1, Т2, Т3, T4; /m3 вытянут в передне-заднем направлении и по длине сопоставим с /m2.
| ___ |