Екатеринбург: УрО РАН, 2009. 100с.
(Смирнов, Большаков Бородин, 1986; Бородин, 1988; Бородин, 1992; Бородин, 1995; Бородин, Давыдова, Елькина, 2006; Бородин, Коурова, Маркова, 2005; Бородин, Елькина, 2006; Галкина, 1977; Зажигин, 1980; Фадеева, 2003a; Фадеева, 2003b; Яковлев, 1994; Яковлев, 1996; Borodin, 1996;и др.)
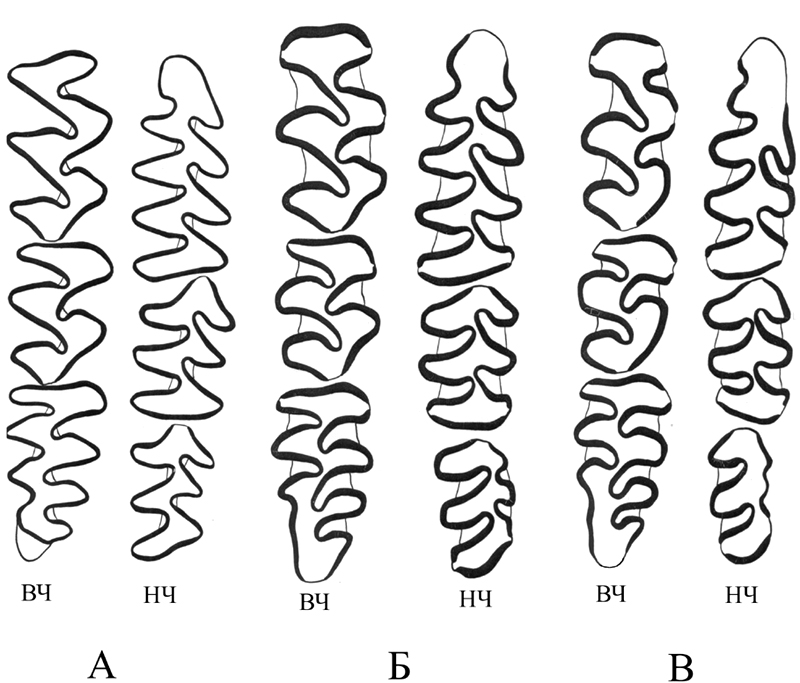
Жевательная поверхность щечных зубов красной полевки. А – молодая особ (корни отсутствуют), Б – взрослая (корни имеются), В – старая особь (высота коронки составляет 1/2 высоты корней).
(min– минимальное значение, m – среднее, max – максимальное значение, n- количество изученных экземпляров; L - длина, W - ширина жевательной поверхности зуба)
|
Признак |
Cl. rutilus (n = 96) |
||
|
min |
m |
max |
|
|
L М1/ |
1.48 |
1.67 |
1.85 |
|
W М1/ |
0.83 |
0.95 |
1.08 |
|
L М2/ |
1.10 |
1.25 |
1.35 |
|
W М2/ |
0.75 |
0.91 |
1.03 |
|
L М3/* |
1.45 |
1.64 |
1.85 |
|
W М3/* |
0.63 |
0.84 |
0.90 |
|
L /m1 |
1.85 |
2.06 |
2.40 |
|
W /m1 |
0.80 |
0.91 |
1.05 |
|
L /m2 |
1.20 |
1.29 |
1.45 |
|
W /m2 |
0.65 |
0.87 |
0.95 |
|
L /m3 * |
1.05 |
1.21 |
1.35 |
|
W /m3 * |
0.60 |
0.73 |
0.78 |
* Количество экземпляров n=30
Положение нижнего резца диагональное: корневые отделы /m1 и /m2 лежат снаружи, а /m3 с лингвальной стороны нижнего резца.
Дентиновые поля призм щечных зубов. Дентиновые поля жевательной поверхности разных зубов слиты(изолированы) в разной степени. У молодых животных грани противолежащих входящих углов смыкаются неполностью.
Отложения наружного цемента во входящих углах плотные, у молодых особей малозаметны..
Эмаль слабо дифференцирована по толщине у молодых экземпляров или дифференцирована по мимомисному типу (более толстая на выпуклых стенках дентиновых призм) и всегда тоньше в вершинах входящих углов.
Дентиновые тракты на передне-буккальной стороне головки непарной петли /m1, задней непарной петле М3/, по обеим сторонам непарных лопастей щечных зубов. На зубах с очень сильно сточенной коронкой дентиновые тракты могут доходить до жевательной поверхности и на других выходящих углах, прежде всего на вершине LSA2 дентинового поля Т1 М1/. У молодых животных эмаль на жевательной поверхности может быть непрерывна.
Корни у щечных зубов формируются. Формирование единой пульпарной полости заканчивается к 2 месяцам, к трем месяцам значительная часть особей имеет замкнутые корни (Тупикова,Сидорова, Коновалова, 1970; Смирнов, Большаков Бородин, 1986). Высота коронки минимальная для рассматриваемых представителей рода.
Строение и морфотипические характеристики жевательной поверхности щечных зубов:
М1/-М2/ без добавочных петель. Полное слияние всех полей жевательной поверхности встречается у наиболее молодых особей и на поздних стадиях стирания коронки , когда могут быть слиты Т3-Т4 на М1/ и Т2-Т3 на М2/.
M1/ – передняя непарная лопасть, Т1, Т2, Т3, Т4;
M2/ – передняя непарная лопасть, Т2, Т3, Т4;
М3/ - передняя непарная лопасть, Т2, Т3, Т4; Т5, задняя непарная петля. В 96,6% случаев передняя непарная лопасть отделена от Т2 и Т3 отделен от Т4 (кроме полностью слитых молодых, составляющих 3,4%), то есть проявляется тенденция к изоляции комплекса Т2-Т3. В 55,2% случаев Т2-Т3 слиты с образованием единого замкнутого пространства. Задняя непарная петля имеет различное строение, встречаются все варианты, описанные для рода. Развитый входящий угол LRA4, доходящий до противоположной стенки зуба, встречен в 17% случаев. 75.9% случаев – LRA4 развит настолько, что формируется четвертый выступающий угол с внутренней стороны, но не доходит до противоположной стенки. У старых особей встречено по одному зубу без LRA4 – в одном из них имелась марка на задней непарной петле, другой был представлен единой округлой задней непарной петлей.
/m1 – задняя непарная лопасть, Т1, Т2, Т3, T4, T5, головка антероконидного отдела как правило скошенно-грибовидной формы. В отличии от рыжей полевки головка антероконидного комплекса массивна и более широко слита с Т5. Варианты слияния дентиновых полей как антероконидного отдела так и основных треугольников могут быть видоспецифичны. Наиболее часто встречающийся вариант строения /m1 – попарное слияние треугольников жевательной поверхности (Т1-Т2, Т3-Т4) и Т5-головка антероконидного комплекса.
/m2 - задняя непарная лопасть, Т1, Т2, Т3, формирующийся Т4. В большинстве случаев (59 из 62) Т1-Т2 и Т3-Т4 слиты попарно, однако в 3 случаях слиты только Т3-Т4.
/m3 – задняя непарная лопасть, Т1, Т2, Т3, формирующийся Т4; Т1-Т2 и Т3-Т4 попарно слиты с образованием единых замкнутых пространств (ромбов) на жевательной поверхности..
Возрастная изменчивость сильно выражена. Как внутри- так и межвидовые сравнения по морфотипическим и размерным характеристикам должны проводиться только в пределах соответствующих возрастных или онтогенетических групп.